文献来源:Lin J, Chen C, Li S, et al. Spatial and single-cell characterization of human glioblastoma tumor microenvironment reveals malignant cellular communities. Nature Neuroscience. 2026; doi:10.1038/s41593-026-02265-5. PMID: 41992007.
研究团队:中国科学技术大学 瞿昆团队

一、研究背景与设计
胶质母细胞瘤(GBM)是成人最常见且致死率最高的原发性恶性脑肿瘤,中位生存期仅约15个月。其高度侵袭性与治疗抵抗性,很大程度上源于肿瘤微环境中复杂且异质的细胞组成与空间排布。传统单细胞测序虽能解析细胞异质性,但缺乏空间定位信息;单纯空间转录组则受分辨率限制,难以精确识别细胞类型。本研究通过整合多组学技术,系统构建了GBM微环境的空间结构全景图谱。
研究纳入100例原发性GBM患者,整合121个组学数据profile,涵盖多个技术平台:
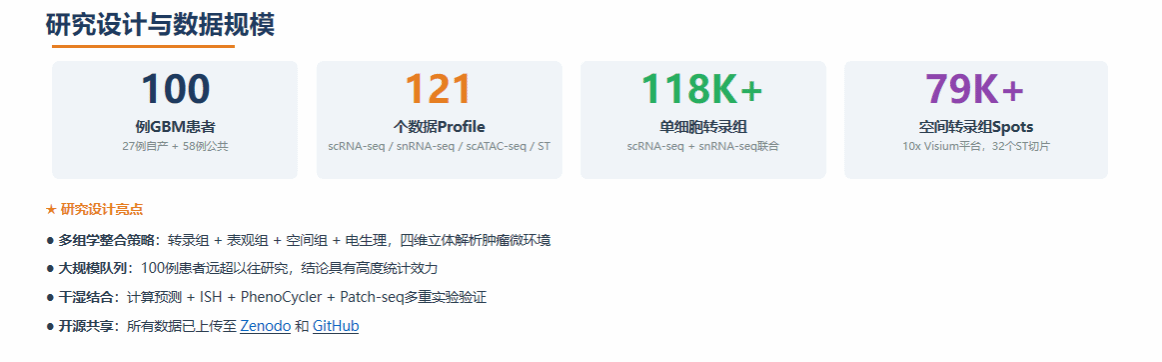

二、核心发现:四大恶性细胞群落(CCs)
研究团队利用自主研发的空间转录组整合分析算法SPACEL(Nature Communications 2023),通过Splane模块联合分析32张ST切片,鉴定出10个空间域(D1–D10)。其中D9、D10为瘤周区域(CNV水平低,富集正常神经元、星形胶质细胞等),D1–D8为恶性区域,进一步聚类为4个在不同患者间保守存在的恶性细胞群落(Cellular Communities, CCs):
上述分类经多轮原位杂交(ISH)在单细胞分辨率下验证,细胞组成与10x Visium数据高度一致。
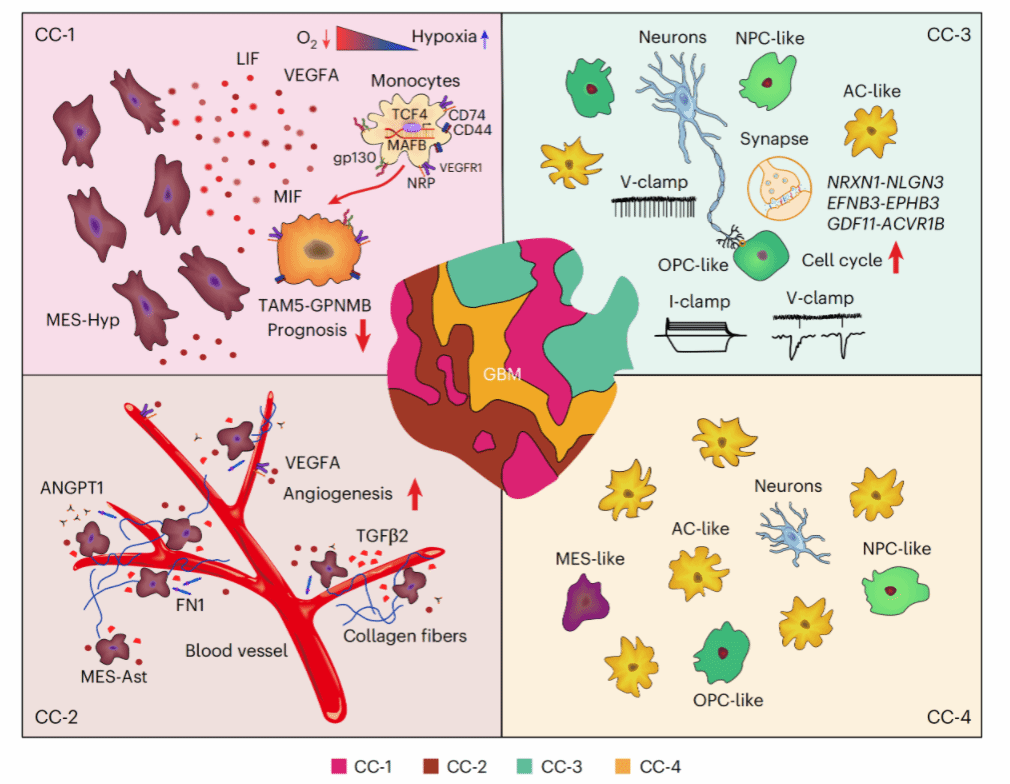
Figure 1研究设计和四大细胞社区的发现
100例患者、121个profiles、4大技术平台整合,鉴定出25种细胞类型,通过cell2location和SPACEL识别10个空间域,最终聚类为4个保守的细胞社区。
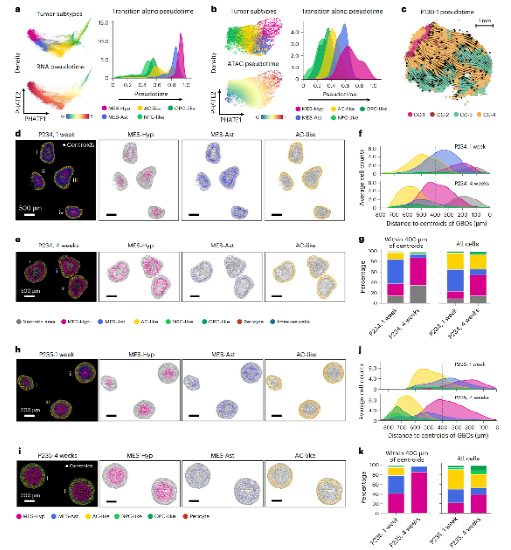
三、MES-like肿瘤细胞的双亚型分化与空间异质性

研究在18,139个MES-like肿瘤细胞中鉴定出两个功能与空间分布显著不同的亚群:
3.1 MES-Hyp(缺氧型)
空间定位:主要富集于CC-1(缺氧核心区)
分子标志:ADM、NDRG1、VEGFA、CA9等缺氧响应基因高表达
细胞互作:与TAM5-GPNMB巨噬细胞显著共定位(P = 4.3×10⁻⁹)
功能机制:通过释放VEGFA等分子诱导TAM表型转变,促进肿瘤发展
3.2 MES-Ast(血管相关型)
空间定位:广泛分布于CC-1、CC-2、CC-4
分子标志:COL1A2、COL1A1、GFAP、LIF、IL11高表达
细胞互作:与内皮细胞/周细胞显著共定位
功能机制:通过释放TGF-β2促进肿瘤血管重塑
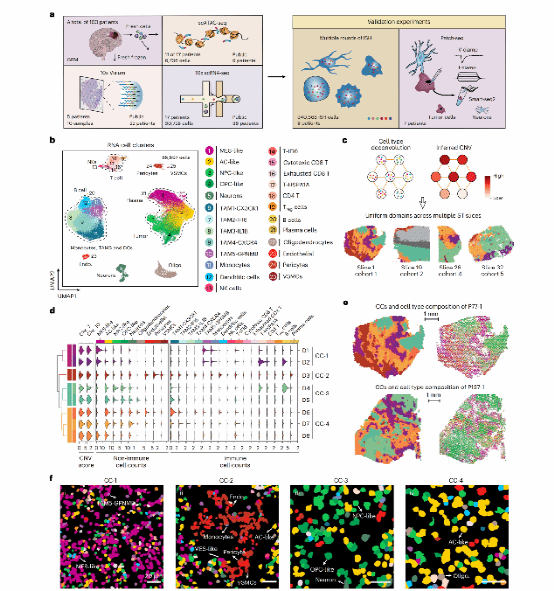
Figure 2:MES-like双亚型:缺氧型 vs 血管型
18,139个MES-like细胞经Leiden聚类分析,鉴定出MES-Hyp(缺氧型)和MES-Ast(血管型)两个功能迥异的亚型。ISH实验验证了两者的空间分布特征。
3.3 亚型转化的动态轨迹
scRNA-seq与scATAC-seq联合拟时序分析揭示:MES-Ast→MES-Hyp→AC-like的转化轨迹,且以MES-Ast向MES-Hyp的单向转化为主。
患者来源GBM类器官(GBO)实验证实,缺氧环境可驱动MES-Ast向MES-Hyp转化,Notch信号在该过程中发挥关键调控作用。
Figure 3 :肿瘤细胞时空动态转化
四、TAM5-GPNMB的极化调控机制
GBM微环境中肿瘤相关巨噬细胞(TAM)占免疫细胞的81.5%。研究将其分为5个亚群,其中TAM1–TAM4高表达稳态小胶质细胞标志基因(TMEM119、CX3CR1),提示来源于小胶质细胞;TAM5-GPNMB高表达单核细胞来源脑巨噬细胞标志基因(CD163、TGFBI)及GPNMB,提示来源于外周血单核细胞。
极化轨迹:通过Palantir算法在转录组与染色质可及性两个层面追踪,发现清晰的单核细胞→TAM5-GPNMB分化轨迹。scATAC-seq分析(6,736细胞,11例患者)鉴定出9个关键转录因子(包括SREBF1、CEBPA、MAFB等),其motif偏差分数在空间转录组数据上与TAM5分布高度吻合。
Figure 4 :TAM5-GPNMB极化调控:单核→巨噬的黑化之路
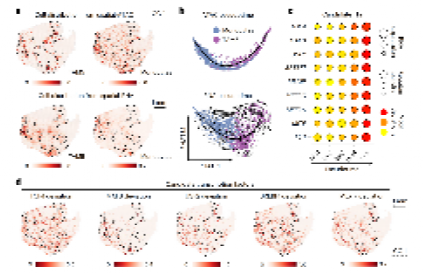
6,736个scATAC-seq细胞经Palantir追踪发现9个关键转录因子(SREBF1, CEBPA等)调控单核→TAM5分化,与M2极化密切相关。GPNMB是TAM5的标志分子。

功能意义:GPNMB高表达的TAM可将髓鞘来源脂质转移到邻近MES样肿瘤细胞,促进肿瘤增殖和复发。TAM5-GPNMB与单核细胞显著富集于CC-1。
五、细胞群落特异的互作网络
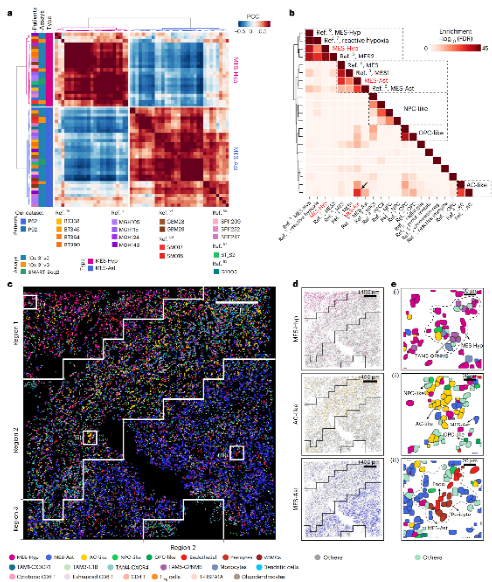
研究整合CellChat配体-受体数据库与COMMOT最优传输算法,结合空间距离约束,推断各CC的细胞间通讯网络,并经PhenoCycler多重免疫荧光实验验证。
Figure 5 :各CC细胞间通讯网络
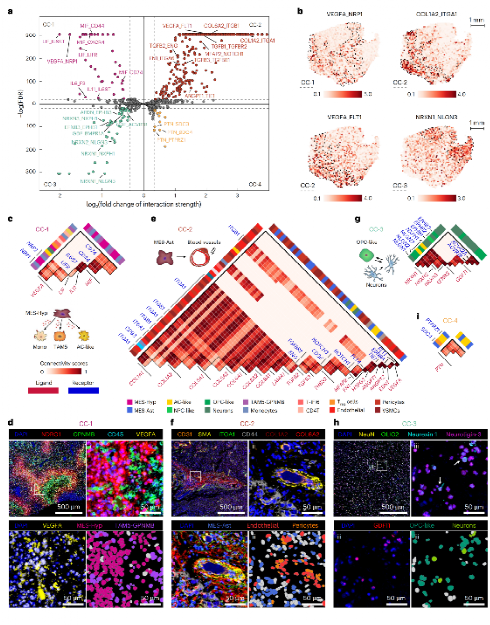
CellChat+COMMOT分析发现每个CC有独特的通讯密码,PhenoCycler 21抗体多重免疫荧光实验验证了关键LR配对。

Figure 6 :Patch-seq验证神经元-OPC-like突触连接
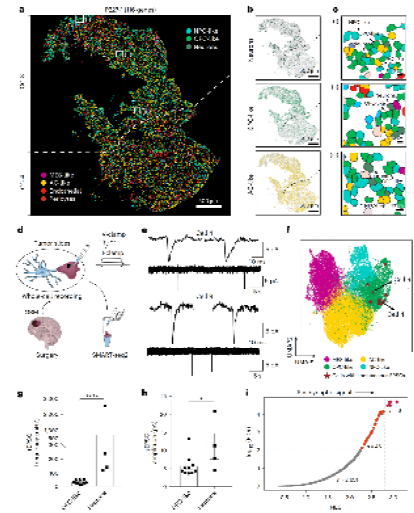
7例GBM患者的21个细胞中,10个sEPSC+细胞全部为OPC-like。GSEA分析证实突触后信号与细胞增殖显著正相关。
神经元与OPC-like肿瘤细胞通过突触黏附分子(NRXN-NLGN家族)及GDF11等信号直接互作。Patch-seq分析(7例患者,21个细胞)发现,10个记录到自发兴奋性突触后电流(sEPSC)的细胞全部为OPC-like肿瘤细胞。GSEA分析证实突触后信号通路与细胞增殖显著正相关。
OPC-like肿瘤细胞高表达PDGFRA和CSPG4,这些基因与突触功能密切相关,提示其继承了正常OPC与神经元互动的社交属性。

图文:陈锦钰
参考文献
[1] Lin J, Chen C, Li S, et al. Spatial and single-cell characterization of human glioblastoma tumor microenvironment reveals malignant cellular communities[J]. Nature Neuroscience, 2026. doi:10.1038/s41593-026-02265-5. PMID: 41992007.
[2] Neftel C, Laffy J, Filbin MG, et al. An integrative model of cellular states, plasticity, and genetics for glioblastoma[J]. Cell, 2019, 178(4): 835-849.e21. doi:10.1016/j.cell.2019.06.024.
[3] Hu J, Li H, Li S, et al. SPACEL: deep learning-based characterization of spatial transcriptome architectures[J]. Nature Communications, 2023, 14: 7603. doi:10.1038/s41467-023-43271-3.
[4] Korsunsky I, Millard N, Fan J, et al. Fast, sensitive and accurate integration of single-cell data with Harmony[J]. Nature Methods, 2019, 16(12): 1289-1296. doi:10.1038/s41592-019-0619-0.
[5] Kleshchevnikov V, Shmatko A, Dann E, et al. Cell2location maps fine-grained cell types in spatial transcriptomics[J]. Nature Biotechnology, 2022, 40(5): 661-671. doi:10.1038/s41587-021-01139-4.
[6] Jin S, Guerrero-Juarez CF, Zhang L, et al. Inference and analysis of cell-cell communication using CellChat[J]. Nature Communications, 2021, 12: 1088. doi:10.1038/s41467-021-21246-9.
[7] Cang Z, Zhao Y, Nie Q. Screening cell–cell communication in spatial transcriptomics via collective optimal transport[J]. Nature Methods, 2023, 20(9): 1326-1335. doi:10.1038/s41592-023-01928-4.
[8] Wang X, Park J, Susztak K, et al. Bulk tissue cell type deconvolution with multi-subject single-cell expression reference[J]. Nature Communications, 2019, 10: 380. doi:10.1038/s41467-018-08023-x.
[9] Biancalani T, Scalia G, Buffoni L, et al. Deep learning and alignment of spatially resolved single-cell transcriptomes with Tangram[J]. Nature Methods, 2021, 18(11): 1352-1362. doi:10.1038/s41592-021-01264-7.
[10] Venkataramani V, Tanev DI, Strahle C, et al. Glutamatergic synaptic input to glioma cells drives brain tumour progression[J]. Nature, 2019, 573(7775): 532-538. doi:10.1038/s41586-019-1564-x.
[11] Venkatesh HS, Morishita W, Geraghty AC, et al. Electrical and synaptic integration of glioma into neural circuits[J]. Nature, 2019, 573(7775): 539-545. doi:10.1038/s41586-019-1563-y.
[12] Osswald M, Jung E, Sahm F, et al. Brain tumour cells interconnect to a functional and resistant network[J]. Nature, 2015, 528(7580): 93-98. doi:10.1038/nature16071.




 地址:上海市徐汇区医学院路138号
地址:上海市徐汇区医学院路138号  邮编:200032
邮编:200032  电话/传真:021-54237056
电话/传真:021-54237056  邮箱:itbr@fudan.edu.cn
邮箱:itbr@fudan.edu.cn 