引言
“在这个社会上,一件简单的事情,把简单的事情重复去做,它就不简单了,但是要坚持。”这是最近很火的网络热梗。虽然大家玩起梗来不亦乐乎,但在笔者看来这其实和我们常说“千锤百炼,百炼成钢”是一样的意思,也可以解读为一个不断的学习过程,在这简单重复的背后蕴藏着不简单的神经机理。我们在不断的学习以及肌肉训练过程中,伴随着大脑当中神经网络的重塑,以使我们更加高效完成任务。这个过程依赖于大脑中两类主要的两类神经元类型(兴奋性的锥体神经元与抑制性的中间神经元)之间连接强度的改变[1]。通常来说,锥体神经元活动模式与学习行为之间存在着很高的相关性[2]。大量研究显示,抑制性中间神经元在学习过程中也表现重要作用[3,4];但由于中间神经元在解剖学和电生理特性中存在极大的异质性,具体每一类中间神经元类型介导的确切作用尚不清楚。
2023年7月20日约翰霍普金斯大学医学院的Hyung-Bae Kwon团队在Nature neuroscience上发表了“An adaptive behavioral control motif mediated by cortical axo-axonic inhibition”的文章。该研究发现中间神经元一类吊灯细胞(ChC)在运动学习过程中,通过对M2运动前神经元中不相关反应的抑制来有效的进行精准的运动控制[5]。

结果
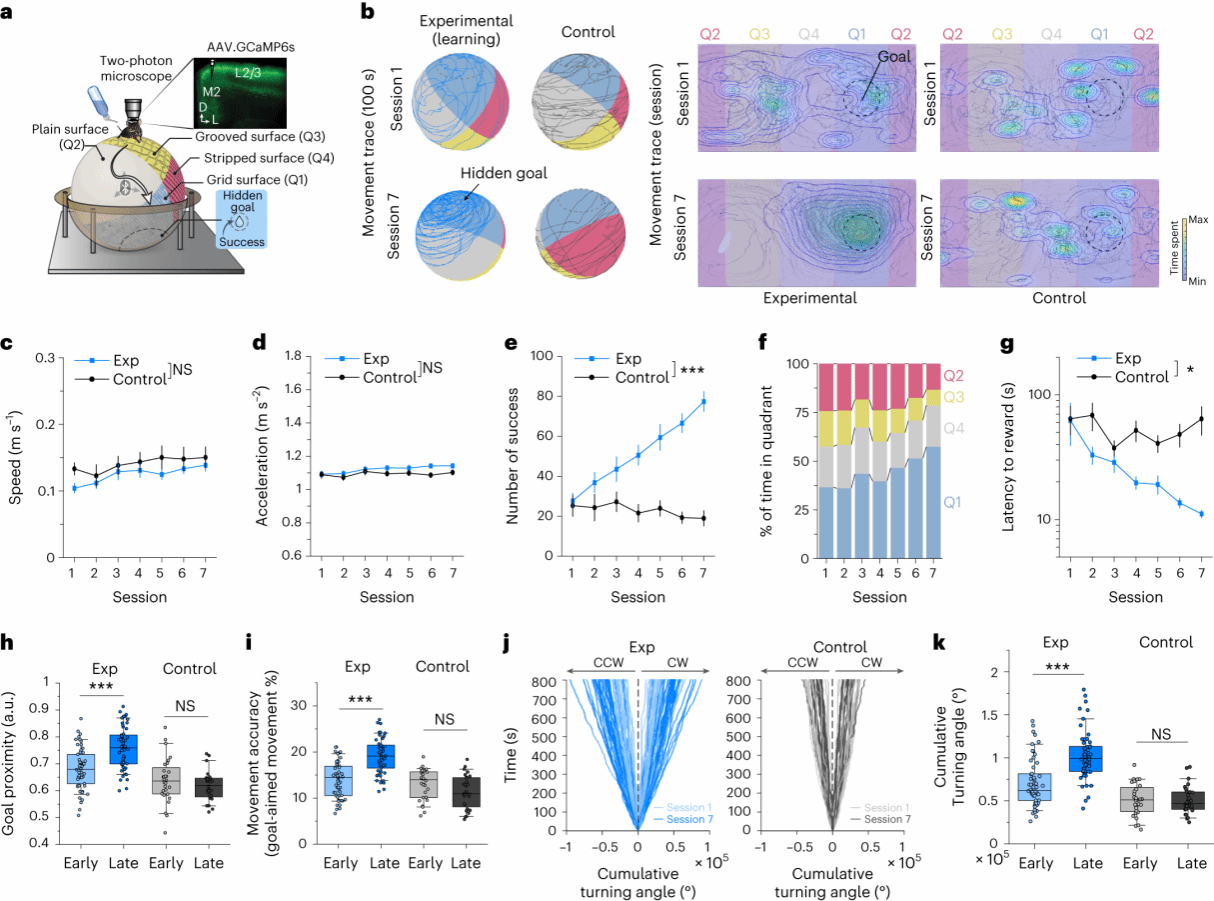
1.运动学习能提高对方向的控制
为了研究M2的运动前神经元在运动学习中的编码规律,作者构建了一个目标导向运动控制的空间导航范式。小鼠被放置于具有四个不同纹理象限的悬浮球上,通过转动悬浮球方向达到目标区域来获得奖励(图1a)。经过连续7次的训练后,禁水小鼠表现出在目标区域停留的时间更长,而未禁水的小鼠并无差异(图1b)。另外,随着训练次数的增加,禁水小鼠获得水奖励的次数也随之增加,获得水的潜伏期越来越短(图1e、f、j)。后期的阶段相较于早期的阶段,其对运动精度以及在目标区域转动的角度也有所增强(图1i)。这些表明,在此运动学习过程中,小鼠对方向控制能力有所增加。
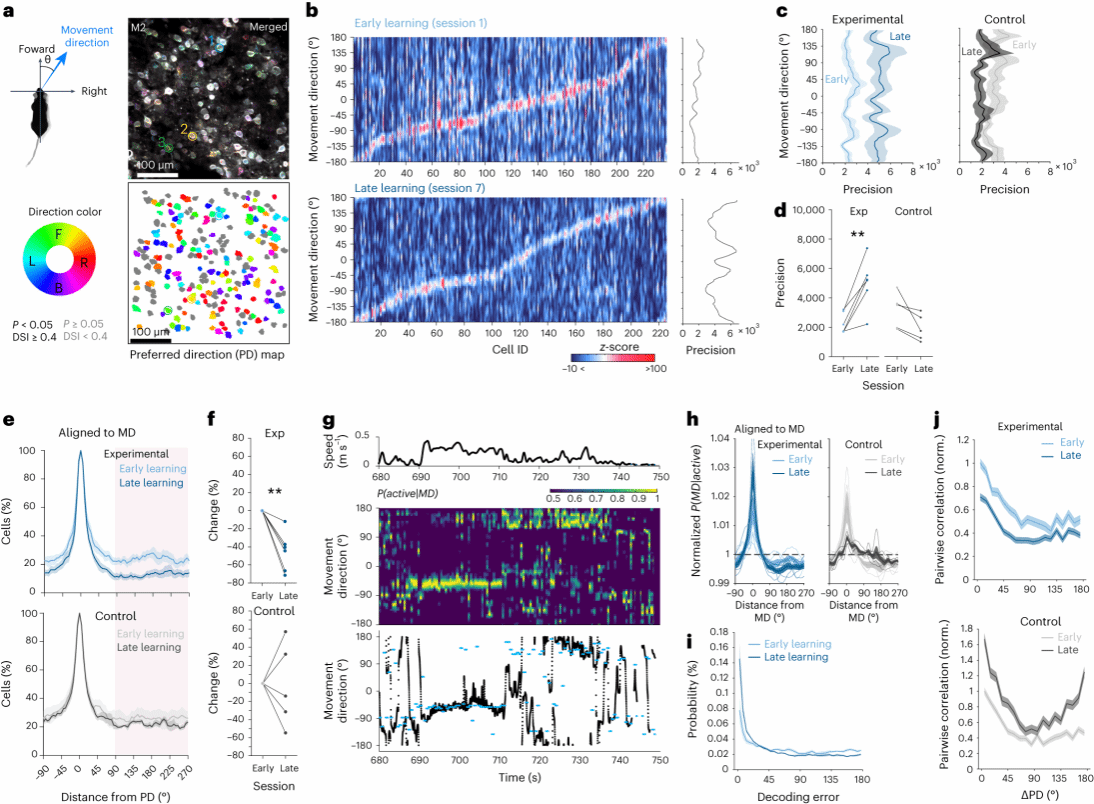
2.运动学习通过抑制不相关方向上的活动来促进M2神经元对方向的精确编码
接下来,作者分析了M2中单个运动前神经元对方向选择性的钙信号反应。不同的神经元群体在其特定运动方向上具有最高的钙活动反应,在到达训练后期,这种对该运动方向的反应精度也有所提高(图2b)。对神经元在运动方向上的活跃程度进行分析,显示当运动方向远离神经元偏好方向时,其活跃的比例随之显著降低,这种现象在学习后期会进一步下降(图2e)。比较偏好方向完全相反两个神经元之间的成对关系发现,在实验组中神经元对的成对相关性随着偏好方向差异的增加而降低,而在对照组中则逐渐增加(图2j)。这些结果表明,不相关方向反应的减少促进了M2神经元对运动方向的精确编码。
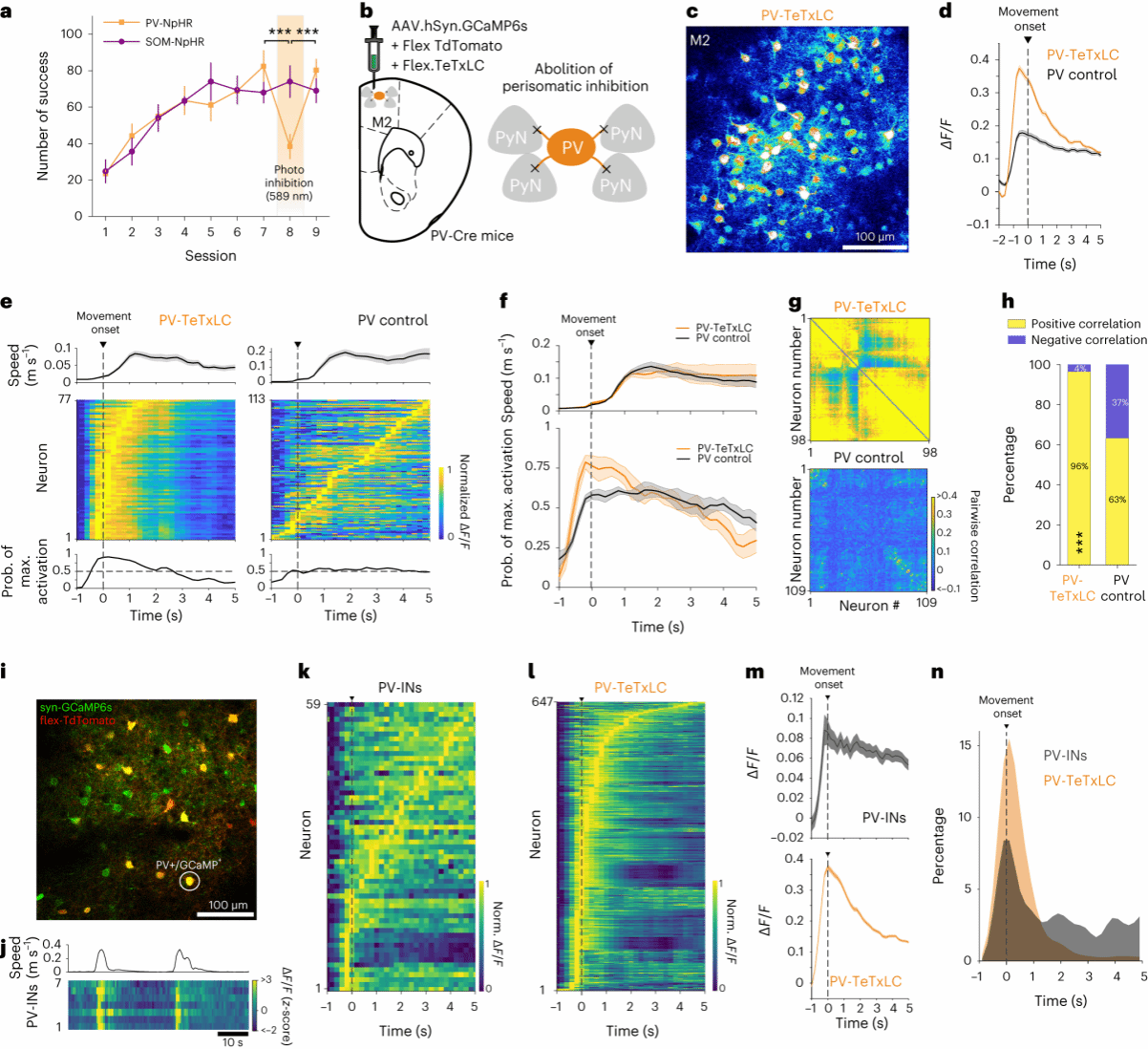
3.损伤PV中间神经元会引起全局兴奋性的改变
那么,学习过程是如何抑制不相关的神经元活动来实现对运动方向进行精确编码的,作者猜测中间神经元可能参与其中的。通过光遗传抑制M2中的PV神经元,可以破坏目标导向的运动表型,而抑制另一类SOM中间神经元,并无影响(图3a)。使用TeTxLC(白喉毒素)病毒特异性破坏PV神经元突触前 GABA释放,观察到运动学习过程中M2神经元的整体活性过度增加,以及在运动开始时M2神经元出现超同步化现象,并在后续运动中神经活动的稀疏性有所降低(图3d、e)。另外,PV-TeTxLC小鼠中M2神经元的成对相关性和正相关的成对比例显著高于PV对照小鼠(图3g、h),相当一部分PV神经元也显示出在运动起始时活动的增加(图3k-n),表明PV-TeTxLC小鼠中M2神经元的过度活跃可能是由于PV神经元对M2神经元抑制反应失效所引起的。这些结果显示,PV神经元对周围M2神经元的抑制参与了任务相关的运动控制,这种抑制性会导致M2神经元全局兴奋性的增强。
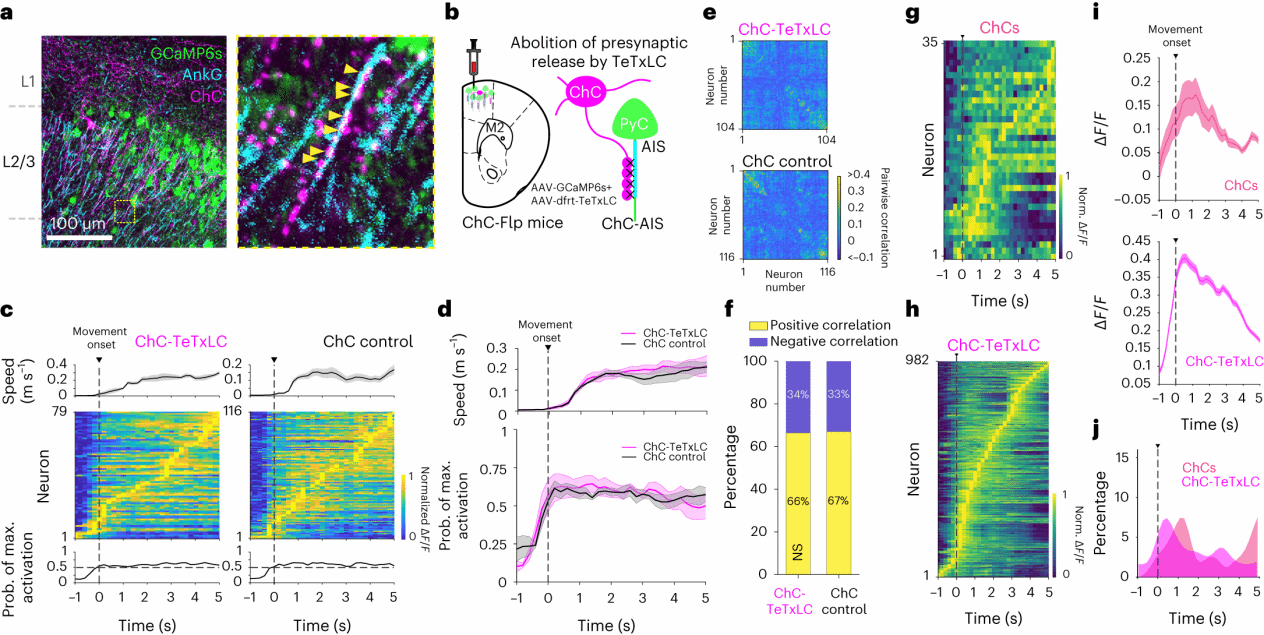
4.损伤ChC不影响对M2神经元的稀疏序列性活动
为了进一步确定对M2神经元抑制的潜在机制时,作者将目光锁定在PV中一类支配锥体神经元轴突起始段(AIS)的 ChC上。 通过在ChC-Flp小鼠中双侧表达FlpO依赖性TeTxLC病毒并观察M2神经元的活性显示,与PV-TeTxLC小鼠的超同步神经元活动相比,并没有观察到全局变化(图4e、g)。相反,在ChC-TeTxLC和对照小鼠的运动过程中观察到稀疏的、连续的活动传播(图4c、d)。成对相关性分析表明两组神经元对之间的局部而非全局相关活动,两组正负相关对的比例也相近(图4f)。与PV-IN相比,大多数ChC并不是运动起始阶段,而是在运动过程中表现出活动增加(图4i),并且 ChC-TeTxLC 不会引起小鼠运动开始时M2神经元的过度活动(图4h、j)。这些结果表明,ChC与PV神经元对M2神经元的调控具有不同的活动特征,沉默ChC的活动不会引起整体网络兴奋性的全局改变。

5.化学遗传学抑制ChC破坏了M2前运动神经元对方向的选择性。
那么,沉默ChC活动是否会影响运动行为任务期间 M2 神经元对方向编码的调整。向 ChC-Flp 小鼠的 M2 中双侧表达 FlpO 依赖性的抑制性化学遗传学病毒(hM4Di),在训练第8个阶段时腹腔给予CNO,会显著损害小鼠目标导向的导航和方向转动能力(图5c、d、e、f)。为了确定ChC在M2神经元方向编码调节的作用,采用同时将FlpO依赖的hM4Di-mCherry和无选择性的GCaMP6s病毒共表达到 M2的L2/3层中。化学遗传学抑制ChC活性,会显著抑制不同的神经元群体在其运动方向上的钙活动反应,对特定运动方向的反应精度变得更差,M2神经元对非偏好方向上的活跃比例有所增加(图5-n)。CNO的使用增加了神经元对之间的成对相关性,而不是在运动学习中看到的神经元对之间去相关的增强(图5o)。这些结果表明,ChC活动的抑制会增加运动行为上无关的神经活动,破环对运动的精确编码。
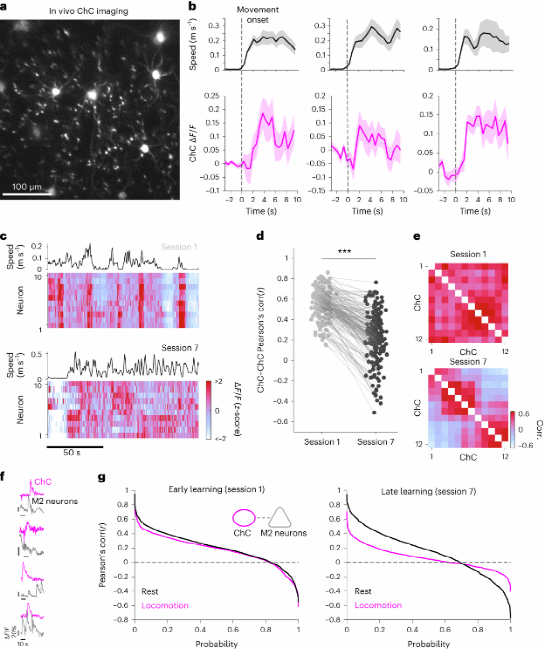
6.运动学习会增加ChC活动的异质性
观察小鼠执行运动任务时ChC活动的动态变化显示,ChC在运动过程中表现出活动增加(图6b)。在训练的第7阶段时,ChC活动具有更高同步性(图6c)。行为任务期间ChC之间的成对相关性降低,ChC的活动变得更加多样化以及非同步化,会形成更多的ChC子簇(图6d)。ChC和相邻的M2神经元之间的活动也显示出随着学习次数增加发生了改变,在运动学习后期阶段,与休息相比,运动期间正相关系数和负系数的总体比例显着降低,表明ChC和M2神经元之间的去相关增强(图6g)。这些结果表明,随着学习的进行,ChC活性变得更加多样,ChC与邻近M2神经元之间群体活动控制的特异性增加。
`
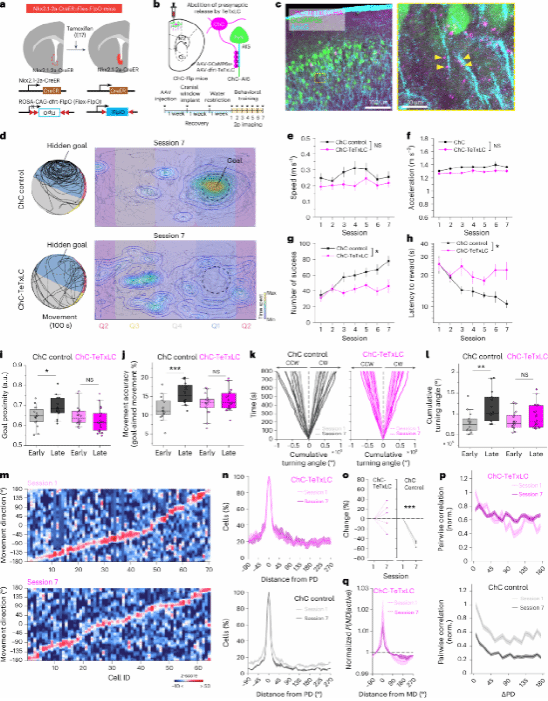
7.损伤ChC功能会破坏运动学习过程中对方向的精确编码
对ChC的损伤是否会影响目标导向任务中的方向编码和运动学习能力呢?同样,使用 TeTxLC来破坏ChC突触前GABA 的释放,ChC-TeTxLC小鼠在学习任务方面表现出缺陷(图7a-l),表明ChC对学习目标导向运动控制的必要作用。对M2神经元在学习任务中钙信号反应的检测,未观察到ChC-TeTxLC小鼠对非偏好方向的活性响应减少以及非偏好方向活性的可能性比例的降低(图7m、n)。此外,在ChC-TeTxLC组中,运动方向的激活概率在学习后期更趋于平缓(图7q),表明由于ChC介导的抑制被阻断后,运动方向编码受损,提示ChC介导了学习过程对不相关方向反应的抑制。这些结果共同表明ChC在提高主动运动控制的分布式神经调节方面发挥了必要作用。
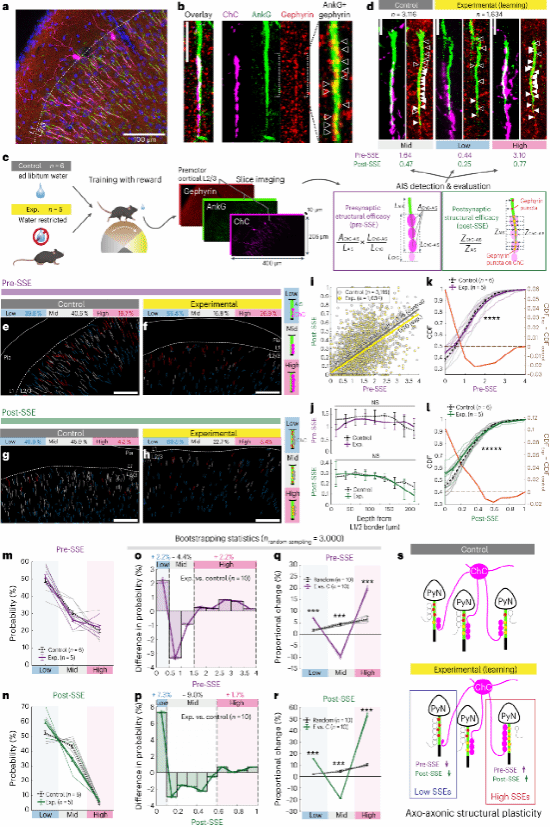
8.运动学习过程中ChC与M2前运动神经元之间轴突-轴突的可塑性发生了改变
在观察分析成像结果时,作者提出了ChC抑制突触强度可能会通过学习发生改变的可能性。于是,通过分析沿AIS的大量抑制性突触前和突触后ChC-AIS结构、ChC轴突button和gephyrin信号(GABA能抑制性突触支架蛋白),分别使用 ChC-AIS 接触和相应的 gephyrin 强度表示突触前结构功效(Pre-SSE)和突触后结构功效(Post-SSE)来估计结构突触强度(图8a-c)。对照组和实验组小鼠中SSE均显示出内在的异质性(图8d-h),表明ChC和M2神经元之间连接的抑制权重不均匀。检测ChC与AIS之间突触异质性的差异是否取决于AIS所处的皮层深度,结果显示整体强度在深度上并没有差异(图8j)。而在经过训练的小鼠,其SSE分布在高部分和低部分的比例有所增加(图q、r)。因此,这些数据表明,ChC神经元在学习过程中对每个轴突突触的抑制进行了重新分配,进而做到有差异的调节。
总结
本研究揭示了前运动皮层(M2)中2/3层的ChC通过抑制不相关的神经元活动来增强运动前神经元对方向调节的编码。另外,突触结构可塑性的定量分析表明,ChC在学习过程中将抑制权重重新分配给单个锥体神经元,做到对单个神经元的选择性抑制,而非对全局抑制。这些结果表明了一个自适应抑制环路的重新分配,能够让ChC精准的控制目标神经元,达到有效的运动控制。
该研究揭示了皮层ChC在经验依赖性可塑性中的特殊抑制作用,这有助灵活性行为的习得。本研究中显示ChC活动的异质性可能已经进化为满足特定功能的需求,而揭示更多神经元类型对神经网络和信号传递的抑制作用,有利于理解认知功能是如何受到内部和外部的精确控制。
原文连接:https://doi.org/10.1038/s41593-023-01380-x
1.Xu, T. et al. Rapid formation and selective stabilization of synapses for enduring motor memories. Nature 462, 915–919 (2009).
2.Peters, A. J., Chen, S. X. & Komiyama, T. Emergence of reproducible spatiotemporal activity during motor learning. Nature 510, 263–267 (2014).
3.Taniguchi, H. et al. A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron 71, 995–1013 (2011).
4.Kvitsiani, D. et al. Distinct behavioural and network correlates of two interneuron types in prefrontal cortex. Nature 498, 363–366 (2013).
5.Jung, K., Chang, M., Steinecke, A. et al. An adaptive behavioral control motif mediated by cortical axo-axonic inhibition. Nat Neurosci 26, 1379–1393 (2023).
本文作者:段树民组赵冰




 地址:上海市徐汇区医学院路138号
地址:上海市徐汇区医学院路138号  邮编:200032
邮编:200032  电话/传真:021-54237056
电话/传真:021-54237056  邮箱:itbr@fudan.edu.cn
邮箱:itbr@fudan.edu.cn 