引言
我们每一天所经历的事情都是在一个特定的时间与空间场景中发生的。时间与空间信息蕴含于我们所经历的所有日常事件之中,比如当你尝试回忆起昨天吃中午饭这件事情时,涉及到你昨天在哪一间餐厅吃饭(空间信息),大概几点去吃的饭以及吃了多久(时间信息)。也就是说,时间与空间构成了我们情景记忆(Episodic Memory)的两个基本维度。因此,研究我们的大脑如何编码,记忆与提取时间与空间信息是神经科学理解人类高级认知功能的关键问题。
内侧颞叶皮层(Medial Temporal Lobe; MTL)是负责情景记忆的一个关键脑区。病灶研究发现MTL受损后病人会出现逆行性遗忘现象,无法回忆起损伤前发生过的事情1。一个关键的问题在于,MTL区域如何编码与整合时间与空间信息?过往很多研究已经揭示了MTL在空间信息编码的重要角色,例如位于海马(Hippocampus)的位置细胞(Place Cells),以及位于内嗅皮层(Medial Entorhinal Cortex; MEC)的网格细胞(Gird Cells)2。以网格细胞为例,当大鼠在一个二维的旷场中自由觅食时,位于内嗅皮层的网格细胞会在大鼠路经该旷场中固定的位置时放电,并且会形成一个稳定的,可以覆盖整个旷场的六边形空间放电模式。这种稳定独特的六边形空间放电模式可以起到在大脑内部形成一个固定的“认知地图(cognitive map)”的作用,指导小鼠的空间导航行为。值得注意的是,当外界环境特征发生改变,位置细胞会发生放电率或者位置偏好的调整,这种现象被称为“remapping”,位置细胞的remapping也被发现伴随着网格细胞整体相位坐标的偏移(realignment)3。由此可见,大脑的空间编码系统可以学习并适应不同空间特征并形成稳定的放电模式来编码空间信息以指导后续行为。确定了大脑存在有处理空间信息的内部机制,研究者进一步想探索我们的大脑是否也存在有类似的机制处理时间信息。
对于时间信息的编码,我们的往往需要处理不同尺度的时长,从毫秒级(例如分辨两声敲击声音的先后顺序)直至数小时级别(例如昼夜节律)。其中,毫秒到数秒尺度的时长对于情景记忆最为关键。对此尺度的时间信息的编码一般被称为Interval Timing。与被广泛研究的MTL如何编码空间信息不同,对于时间信息与MTL的关系目前的研究还相对不足。先前有工作在MEC中发现了一种特定的细胞类型,该细胞在神经元群体的层面上在一段时间间隔内每一个神经元在特定的时间点依次放电,通过形成一个神经序列(Neural Sequence)以编码该时间间隔。这种细胞类型被称之为时间细胞(Time Cell)4。一个问题是,MEC细胞对时间信息的处理是否有着与处理空间信息类似,甚至说是共同的神经机制?同时,我们日常生活中需要面对具有多种空间特征的空间信息,同理,在时间层面,我们也需要处理具有多种时间结构(temporal structure)的场景(context)(尝试想象一下,你在两个不同的餐厅点同一道菜,你在餐厅A可能需要等待5分钟,而餐厅B的厨师可能倾向于更加“慢工出细活”,你可能需要等待15分钟才能吃到这个菜,那么下一次当你挑选餐厅时,你只有掌握了这两所餐厅的这个时间信息你才能做出更好的选择)。那么,面对不同时间结构的场景,我们的大脑如何灵活编码多种时间结构特征并指导我们的行为呢?
今年六月,美国犹他大学神经生物学系的James Heys实验室发表在Nature Neuroscience的工作为上述问题提供了实验性的解释5。该工作通过小鼠行为学,钙成像技术以及化学遗传等工具说明了,MEC时间细胞通过形成情景依赖性(context-dependent)的神经序列活动,或者说情景独特的“时间线(Time Line)”来编码具有不同时间特征的情景信息。
一,小鼠通过训练学习在具有不同时间特征的环境下反应
为了观察小鼠在不同时间结构特点环境下的反应,作者首先根据经典的工作记忆Delay Nonmatch To Sample(DNMS)任务设计了一个具有时间信息加工要求的temporal DNMS任务(tDNMS)。具体来讲,在tDNMS中,头部固定并且禁水后小鼠接受先后两个呈现时长不同(2s或者5s)的气味刺激,两个刺激之间间隔3s。头部固定的小鼠被放置于一个圆柱跑步机上,小鼠四肢可以自由活动驱动底下的跑步机。试次之间间隔(ITI)16-24s。根据两个不同时长的刺激,实验分为三个试次类型:“短-短(S-S)”,“短-长(S-L)”,以及“长-短(L-S)”,即“刺激匹配match型(两个都是短的)”或者“刺激不匹配nonmatch型(一长一短)”。训练规则为经典Go/No-Go条件:试次若是nonmatch型,小鼠需要在第二个刺激之后3s内舔舐获得饮水奖励;若是match型,小鼠则需要抑制舔舐行为(图一)。
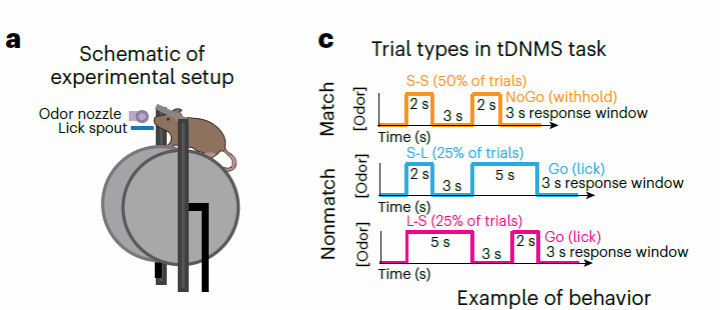
图一,tDNMS任务范式
训练分为两个步骤进行(图二);研究者先训练小鼠学习在两个nonmatch的气味刺激后去舔舐而不能在第一个刺激后或者刺激间隔时舔(pretrain shaping阶段);随后第二阶段研究者进一步引入match试次类型训练小鼠区分两种实验条件,根据每轮试次的时间信息特征选择性地舔舐或者抑制行为(tDNMS阶段)。
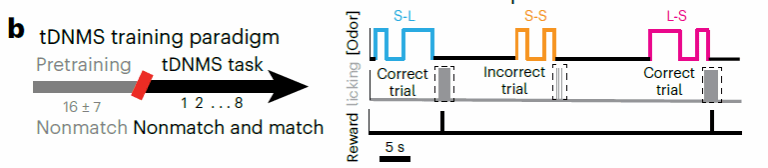
图二,TDNMS训练流程
随着训练次数的增加,小鼠的正确率逐步上升(图三)。在不同试次类型中,在tDNMS阶段里,由于经过了预训练阶段,小鼠在“短-长”类型保持着较高的正确率;引入了match类型后,随着训练次数增加,在“短-短”型试次的正确率也逐步上升;值得注意的是,小鼠在“长-短”类型试次中仅有大约56%的正确率,低于预训练环节将近86.3%的正确水准。研究者发现,“长-短”类型试次中错误的试次主要是由于提前舔舐的冲动行为造成的,这在某种程度上说明了小鼠熟练学习了不同试次的时间结构信息(如果先呈现了“长”刺激,那么该试次肯定要求舔舐行为而非抑制,小鼠有可能会提前进行舔舐行为)。综上,小鼠可以较好地被训练掌握了tDNMS任务的基本要求。
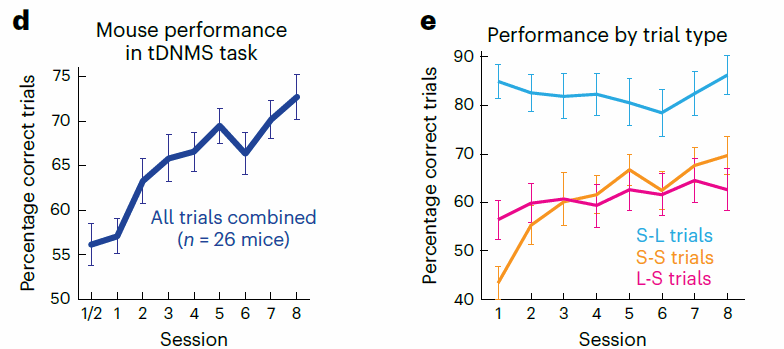
图三,小鼠任务训练学习曲线
二,MEC时间细胞呈现出不同时间特征情景下特异的神经活动
接下来研究者进一步采用细胞分辨率的双光子钙成像技术观察MEC的神经元活动。在所记录到的2056个神经元中,大约33%的神经元表现出规律的时间锁定(Time-locked)活动。在这些神经元中,大约有一半以上的神经元表现出稳定的放电活动模式(放电模式在三种试次类型中有着稳定的特征,例如图四Time Cell 1);除此以外的神经元展现出了情景特异性的放电模式,例如放电模式在某些试次里的消失(Time Cell 3-5),调节放电率大小(Time Cell 6),或者是峰值潜伏期变化(Time Cell 2)。
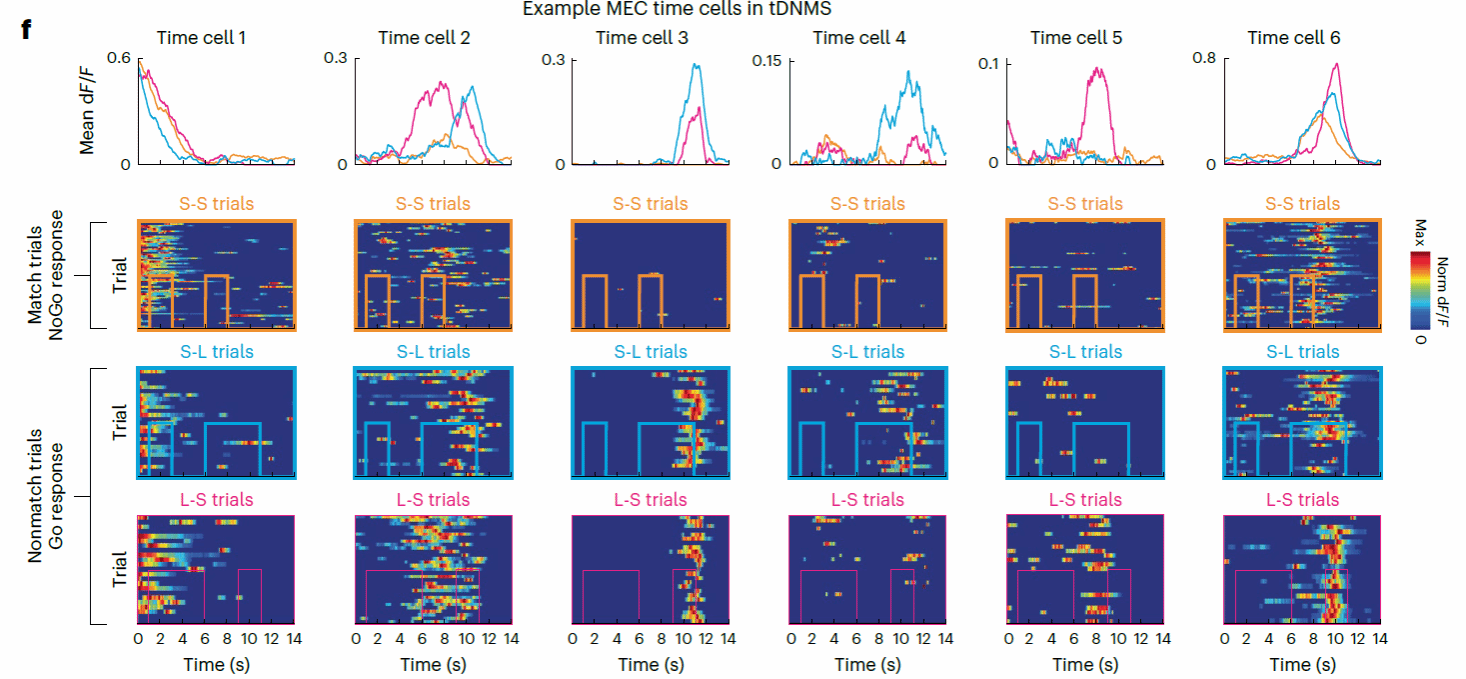
图四,单神经元放电模式
进一步观察神经元群体放电活动规律,研究者发现MEC细胞在每个试次内表现出典型的神经序列放电模式(图五)(神经元集群内每个神经元在特定时间点依次放电,形成一个具有随时间演进的神经序列性放电模式,下图为每个试次类型中按照单细胞神经活动峰值潜伏期从小到大排列后的可视化结果)。
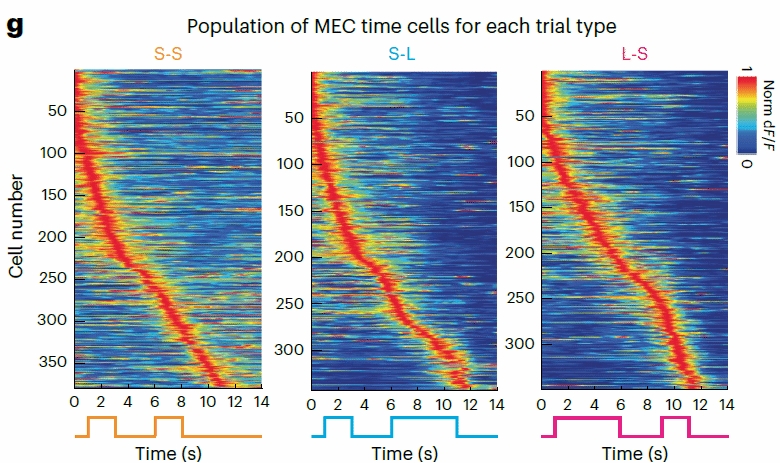
图五,情景特异的神经活动序列
由于过往研究证实MEC在空间信息编码的重要作用,研究者进一步检验了时间信息(试次持续时间)以及空间信息(圆柱跑步机所记录的小鼠活动距离)对MEC神经活动的贡献大小。
通过计算以时间或者距离为参考的单神经元神经活动的变异系数(CV),研究者发现以时间作为参考的MEC神经元整体变异系数小于以距离作为参考所得出的变异系数(图六)。说明该任务中MEC神经元主要响应于时间信息。同时研究者也进一步通过广义线性模型(General Linear Model;GLM)方法量化了时间与距离相对于神经活动的贡献,结果同样支持时间信息的贡献高于距离信息。
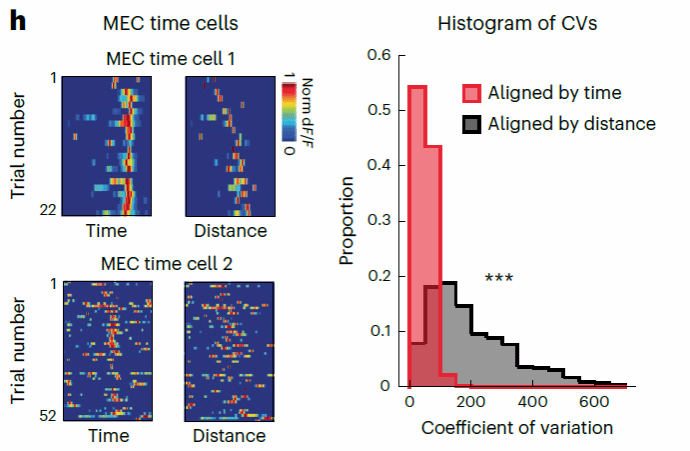
图六,时间细胞活动变异系数分布
综上所述,以上结果标明在执行tDNMS任务时,MEC神经元收到时间信息的调控,并且展现出了情景依赖性的放电模式,在群体神经元水平上表现为特异性的神经序列活动。
三,情景特异的神经序列的形成促进小鼠tDNMS任务的学习
研究者进一步追问这种神经序列的形成是否与小鼠的学习(对tDNMS任务熟练掌握)过程有关。如果相关,那么可以预测的是,(1)这种特异性的神经序列随着训练过程逐步产生;(2)在错误试次中,这种序列模式会被破坏。
为了检验第一个假设,研究者计算MEC时间细胞平均神经元活动在Match vs Nonmatch试次之间的相关性(相关性越低,说明在不同时间特征情景中MEC所产生的序列更加不同)。研究者对比了tDNMS任务训练第一天(Day 1)以及Day N(正确率大于70%的第一天)的细胞活动的相关系数,结果发现在Day N 中时间细胞的Match vs Nonmatch 放电率相关系数更低(图七),这说明,随着训练程度提高,不同时间特征情景下的时间细胞神经序列活动更加具有区分度。

图七,时间细胞神经序列随着训练次数的变化
研究者进一步通过对比时间细胞在不同试次类型之间放电率的差别(图八d-f),并使用线性判别模型(Linear Discriminative Model; LDM)和支持向量机(Support Vector Machine; SVM)对比Day1 和 Day N时间细胞群体神经活动解码试次类型的准确性(下图g-h)。以上分析结果进一步说明,随着小鼠训练学习程度上升,MEC时间细胞神经活动序列对不同时间特征环境的情景依赖性更强,不同试次类型之间的时间细胞活动区别性更大,这符合上述的假设一的推论。
为了验证上述的推论二,研究者比较了错误与正确tDNMS试次之间MEC时间细胞活动的差异。研究者通过对比正确试次vs正确试次时间细胞活动相关系数以及正确试次vs错误试次之间的相关系数发现,MEC时间细胞在正确试次之间展现出比正确与错误试次之间更高的相关性(下图I-J),同时,这种在Day N的正确试次vs错误试次相关性显著低于Day 1的相关性。以上结论符合推论二的假设,当小鼠未能正确完成tDNMS任务,所观察到的情景依赖性MEC神经活动序列模式会损害。
综上所述,研究者通过多种数据分析方法验证了在tDNMS任务中所观察到的MEC时间细胞神经序列活动随着学习程度上升情景依赖性更强,并且可以指导小鼠正确完成tDNMS任务。
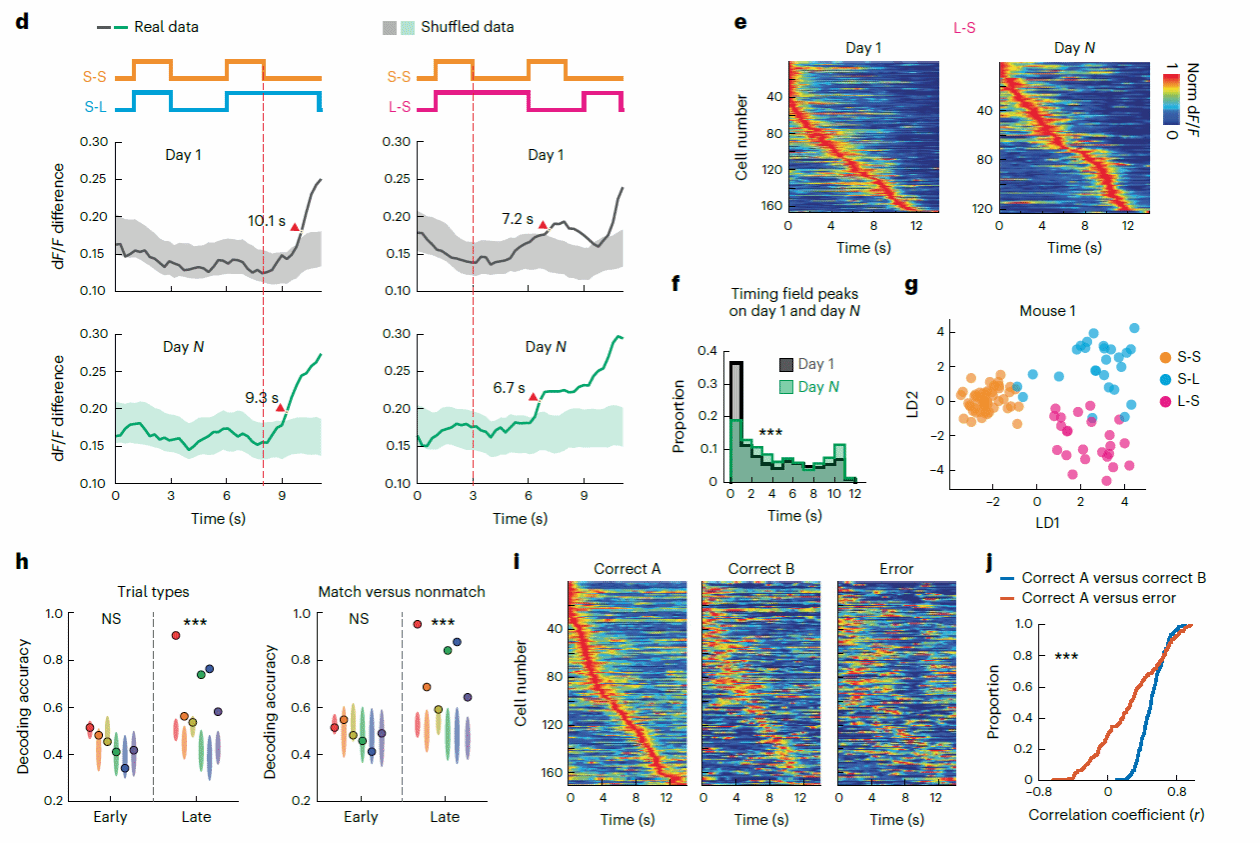
图八,神经活动解码分析
四,MEC仅为学习时间情景依赖性行为所必须
研究者接下来通过化学遗传方法研究特异性地抑制MEC神经元活动是否会影响小鼠tDNMS的学习过程.在小鼠接受tDNMS任务训练前,研究者在双侧MEC注射表达hMD4的AAV病毒特异性抑制MEC神经元活动.结果显示,与控制组相比,接受了化学遗传的小鼠无法成功学习tDNMS任务(图九).特别地,在Match类型试次上,经过8轮训练后接受了化学遗传的小鼠只能达到约50%的正确率.以上结果说明了MEC对学习tDNMS任务所需要的时间情景依赖性行为的必要性.
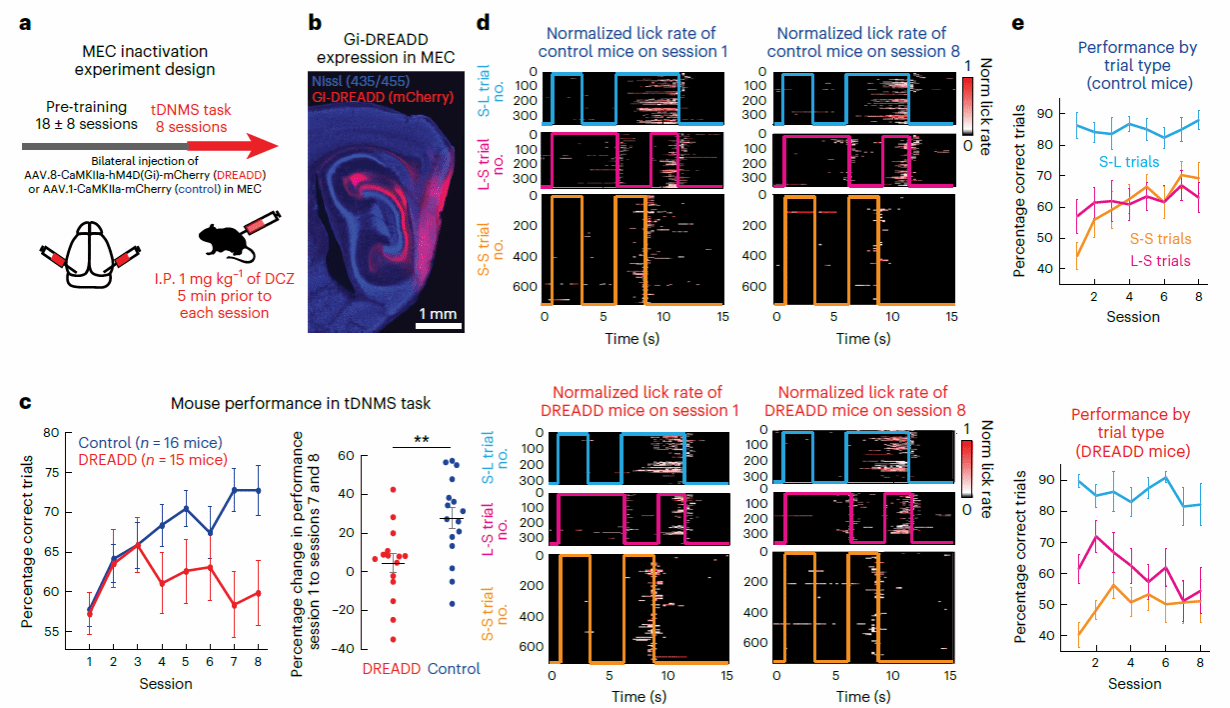
图九,化学遗传干预实验
但是,有趣的是,当研究者在小鼠完成训练后再进行化学遗传干预实验发现,控制组和实验组小鼠的正确率均没有显著下降.这说明,MEC仅对时间情景依赖的学习过程起作用,当这种行为模式学习完成后,小鼠的时间情景依赖行为便不再依赖于MEC.
同时,研究者也发现抑制MEC活动不影响小鼠学习有规律的时间性行为.当训练实验组和控制组小鼠共同完成一个规律性的时间学习任务时(训练小鼠每10s舔舐一次获得饮水奖励),两组小鼠可以被训练达到类似的准确率.以上结果说明,MEC参与有不同时间特征场景调节需求的灵活性行为的学习,而学习固定规律性的时间行为不需要MEC.
五,小鼠使用特定策略完成tDNMS任务
为了完成tDNMS任务,小鼠可能通过以下集中策略完成: (1)以长刺激作为线索,识别每个试次中是否存在有长的刺激,并且在第二次刺激后反应;(2)计算整个试次的时长,对时长更长的试次作处舔舐反应;(3)以长刺激作为线索,但同时计算两个刺激的时长;(4)计算两个刺激的时长并进行比较.研究者称以上四种策略为“策略指导的行为(strategy-based)”
与此同时,由于实验数据显示小鼠并不是100%正确率完成任务,小鼠也可能仅仅依赖不同时长的气味刺激作为线索提示改变每次舔舐的概率,而不用学习三种试次的整体刺激呈现结构。研究者称这种方式为“线索指导行为(cue-based)”。为了研究小鼠采取上述那种行为模式完成tDNMS任务,研究者进一步通过泊松过程建模对实验数据进行拟合。
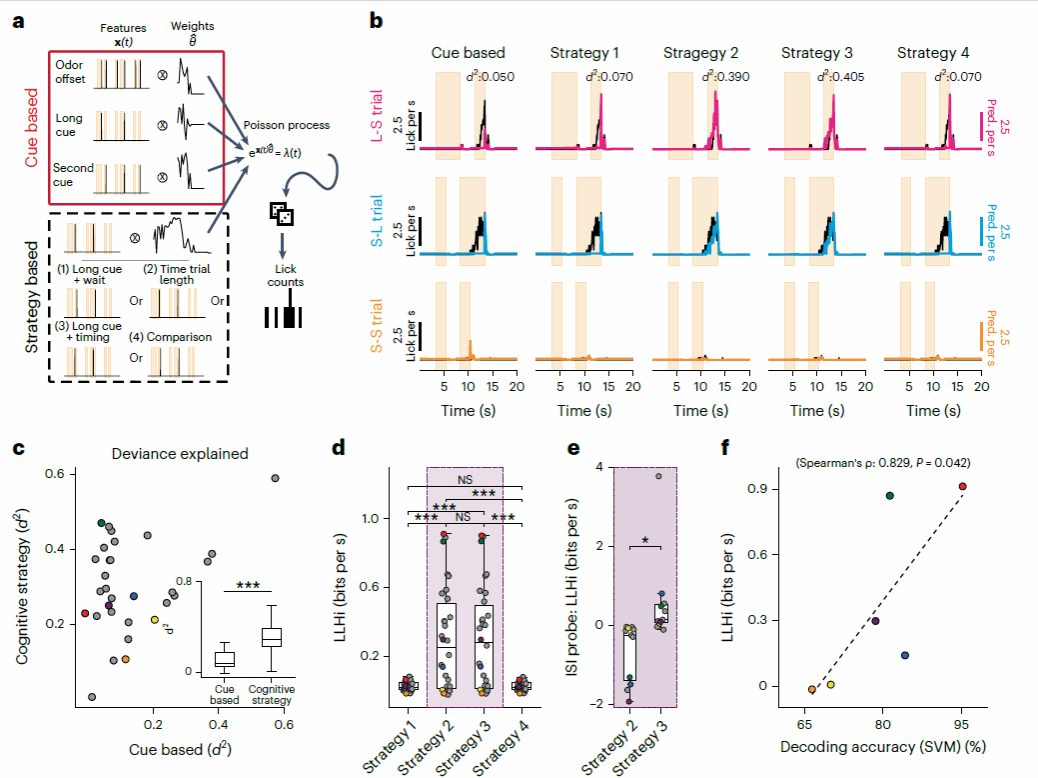
图十,泊松过程建模
建模结果提示,基于“策略指导行为”的拟合结果显著优于基于“线索指导行为”的模式(图十 a-c)。在四种策略中,模型拟合度提示策略2与策略3显著优于策略1和4.为了确定小鼠更可能采取策略2还是策略3.研究者在tDNMS任务中设置了一部分“Probe Trials”(见图十一),其刺激间间隔时间(ISI)为5s,长于标准ISI(3s)。
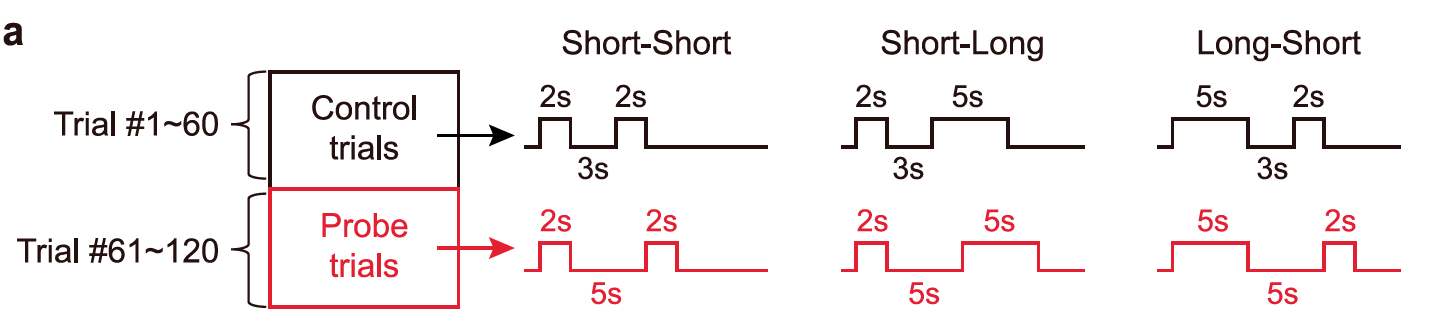
图十一,tDNMS probe trials结构
研究者使用标准试次作为模型训练集并将模型在Probe Trials中进行测试,结果显示策略3的拟合优度显著高于策略2(图十 e),并且其拟合优度与SVM解码准确性显著相关。这说明小鼠最有可能通过采取策略2:以长时间刺激作为线索提示并且对整体试次长度进行计时,来完成tDNMS任务,并且这种策略的使用与MEC活动紧密相关(图十 f)。
六,MEC网格细胞和时间细胞可能共享同一个神经计算机制
过往对MEC网格细胞的研究发现,网格细胞动力学可能是通过连续吸引子网络模型(Continuous Attractor Network; CAN)驱动的6。在CAN网络中,通过在神经元集群中形成局部的循环兴奋性连接,可以在局部神经元群体内部产生活动波包(Activity Bump)。
如果MEC时间细胞与网格细胞同样由CAN网络动力学驱动,那么一个推论是:时间细胞的相对相位活动应该是相对一致的。为了检验以上推论,研究者计算了时间细胞在tDNMS任务中正式试次以及试次间隔(ITI)期间神经元之间的活动相关性(图十二a)。结果显示,神经元之间的活动相关性在正式试次与ITI期间呈现显著的正相关关系(图十二 b),这种相关性在不同试次类型以及错误试次中均保持稳定。并且,在tDNMS任务中,相近时间点放电的时间细胞,即拥有相似相位活动的细胞,更倾向于在试次间隔期间共同放电(图十二 c)。
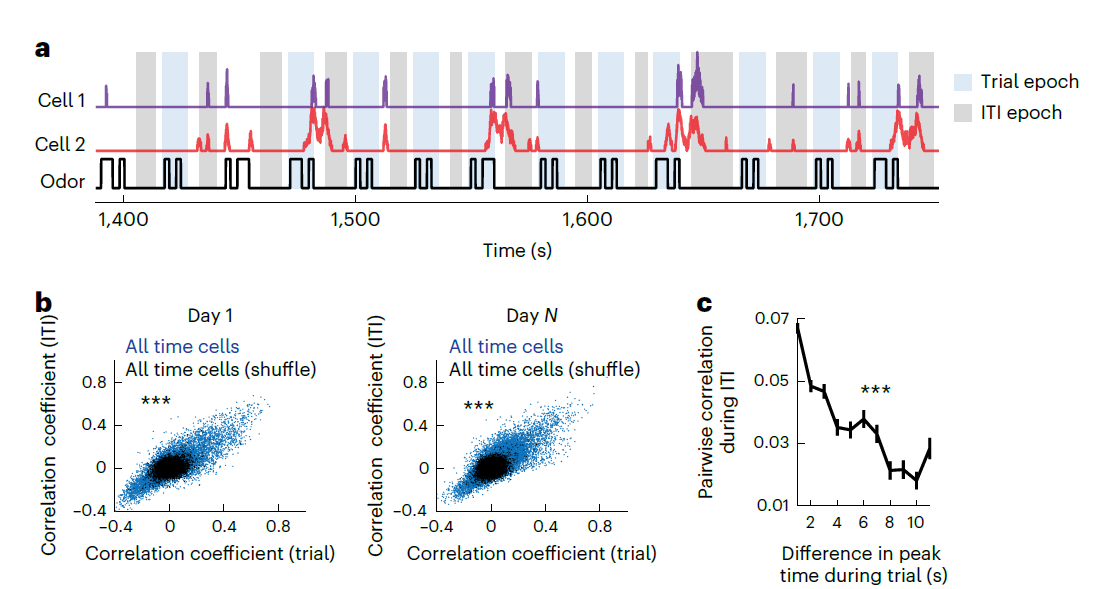
图十二,时间细胞活动相位协同性分析
以上结果说明,MEC时间细胞具有规律的序列放电模式,符合CAN神经动力学的基本假设,有可能与网格空间细胞共享同一套神经环路和计算机制。
总结
综上所述,这篇研究证明了在具有多种时间信息特征的场景中灵活执行行为任务时,MEC通过形成情景特异的神经序列活动支持这种情景间行为转换的学习过程。这种时间细胞神经序列活动随着任务学习过程而逐步产生,并且这种情景特异的神经序列仅仅与本研究中涉及的多种时间特征情景间行为灵活处理有关,完成一个规律性固定的计时任务或者重现复刻已学习好的时间结构信息并不需要MEC的参与。同时,MEC时间细胞可能与网格细胞共享同一套神经机制,进一步验证了时间与空间信息在神经系统编码与存储上具有高度相似性。
未来的工作可以进一步研究其他与时间信息处理相关的脑区,例如海马,外嗅皮层等,在执行如tDNMS需要对不同时间信息特征场景进行灵活行为转换的任务时的角色。
参考文献
1. Tulving, E. Memory and consciousness. Canadian Psychology / Psychologie canadienne 26, 1–12 (1985).
2. Rowland, D. C., Roudi, Y., Moser, M.-B. & Moser, E. I. Ten Years of Grid Cells. Annual Review of Neuroscience 39, 19–40 (2016).
3. Fyhn, M., Hafting, T., Treves, A., Moser, M.-B. & Moser, E. I. Hippocampal remapping and grid realignment in entorhinal cortex. Nature 446, 190–194 (2007).
4. Heys, J. G. & Dombeck, D. A. Evidence for a subcircuit in medial entorhinal cortex representing elapsed time during immobility. Nat Neurosci 21, 1574–1582 (2018).
5. Bigus, E. R., Lee, H.-W., Bowler, J. C., Shi, J. & Heys, J. G. Medial entorhinal cortex mediates learning of context-dependent interval timing behavior. Nat Neurosci 1–12 (2024) doi:10.1038/s41593-024-01683-7.
6. Giocomo, L. M., Moser, M.-B. & Moser, E. I. Computational Models of Grid Cells. Neuron 71, 589–603 (2011).
本文作者:周尚林组-何华葳




 地址:上海市徐汇区医学院路138号
地址:上海市徐汇区医学院路138号  邮编:200032
邮编:200032  电话/传真:021-54237056
电话/传真:021-54237056  邮箱:itbr@fudan.edu.cn
邮箱:itbr@fudan.edu.cn 